遗传发育所等发现植物转亚硝基化酶
一氧化氮(nitricoxide,NO)是一种高度保守的信号分子,参与调控众多生物学过程。NO通过对靶蛋白特异半胱氨酸残基进行S-亚硝基化修饰(S-nitrosylation)是其发挥生物学功能的主要方式之一。与其它蛋白质翻译后修饰不同,...
一氧化氮
(nitric oxide, NO)
是一种高度保守的信号分子,参与调控众多生物学过程。
NO
通过对靶蛋白特异半胱氨酸残基进行
S
-
亚硝基化修饰
(
S
-nitrosylation)
是其发挥生物学功能的主要方式之一。与其它蛋白质翻译后修饰不同,亚硝基化修饰曾被认为是一个非酶促反应,其特异性主要决定于
NO
的局部浓度和靶蛋白的结构。近年来的研究发现
S
-
亚硝基化修饰选择性
(selectivity)
的一种新机制,即一类蛋白可以将其携带的
NO
基团传递至另一个蛋白,导致后者的亚硝基化修饰,这一过程被称为转亚硝基化修饰
(transnitrosylation)
,而介导转亚硝基化的蛋白被称为转亚硝基化酶
(transnitrosylase)
。目前,在动物和大肠杆菌已经发现了数个结构迥异的转亚硝基化酶。在植物中,尚未发现转亚硝基化酶。
中国科学院遗传与发育生物学研究所植物基因组学国家重点实验室左建儒研究组与合作者最近发现了植物特异的转亚硝基化酶。
NO
的主要生物学活性形式是亚硝基谷胱甘肽
(
S
-nitrosoglutathione, GSNO)
,可被高度保守的
GSNO
还原酶
(GSNO reductase, GSNOR)
不可逆分解。因此,
GSNOR
是调控
NO
动态平衡的主控因子,不同物种中
gsnor
突变导致
NO
水平升高以及各种严重的缺陷。通过遗传筛选,研究团队获得了拟南芥
gsnor1
突变体的抑制子突变
rog1
(
repressor of gsnor1
)
。除抑制
gsnor1
突变体的表型外,
rog1
突变体对
NO
的敏感性显著降低,表明
ROG1
是调控
NO
信号通路的一个重要组分。分子遗传学和生物化学研究发现
ROG1
编码一个转亚硝基化酶,其底物之一为
GSNOR1
本身。
ROG1
介导
GSNOR1
的亚硝基化修饰导致其通过自噬途径被降解,从而形成一个正反馈环调控
NO
信号通路。
令人吃惊的是
ROG1
即为过氧化氢酶
CAT3 (catalase 3)
。
ROG1
转亚硝基化酶仅具有很低的过氧化氢酶活性;与之相反,其同源蛋白
CAT2
具有很高的过氧化氢酶活性但很低的转亚硝基化酶活性。决定
ROG1
与
CAT2
酶活特异性的一个主要因素是其各自特异的
Cys-343
残基和
Thr-343
残基。将
ROG1
的
Cys-343
替换为
Thr
后,其转亚硝基化酶活性显著降低,而过氧化氢酶活性升高;反之,将
CAT2
中的
Thr-343
替换为
Cys
后,其转亚硝基化酶活性升高,而过氧化氢酶活性降低。对水稻
ROG1-like
蛋白
(OsCATA)
和
CAT2-like
蛋白
(OsCATC)
的特异性酶活分析得出相似结论,表明这是植物中一种高度保守的机制。该研究发现了调控植物特异选择性
S
-
亚硝基化修饰的
新机制。
上述研究由左建儒研究组与周俭民研究组、李家洋研究组、河南大学宋纯鹏研究组合作完成。相关结果于
4
月
23
日在
Developmental Cell
在线发表
DOI:10.1016/j.devcel.2020.03.020
)
。左建儒研究组博士后陈立超为论文的第一作者,左建儒为通讯作者。该项研究得到国家自然科学基金委、中科院、植物基因组学国家重点实验室的资助。
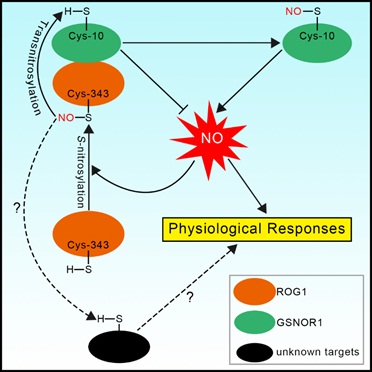
ROG1
介导的一氧化氮信号转导模式图